Estrogen’s role in the induction of parturition and fetal maturation in periparturient ewes
(AS2100-13, December 2023)The objective of this study was to obtain a better picture of estrogen’s role in the control of parturition and fetal maturation in periparturient Rambouillet ewes through the evaluation of the relationship between systemic estradiol (E2) levels and 1) the timing of parturition, lamb’s birth weight, and lamb vigor, and 2) the uterine progestogenic responsiveness and progesterone (P4) plasma levels. Results suggest that estradiol may downregulate myometrial P4 receptor protein expression, leading to myometrial contraction-inducing parturition; however, it does not seem to affect lamb birth weight or vigor.
Results and Discussion
In experiment 1, the hours from treatment to delivery were less in E compared with the C group (64.1 ± 75.64 vs 374.4 ± 75.64 h, P = 0.01). This indicates that the E group had an average day of gestation (DOG) at lambing of approximately d 142 ± 3, while the C group had an average DOG of approximately d 154 ± 3. In a more recent study (Davila-Ruiz, Reynolds, Conley et al., unpublished) we found with that a similar treatment with E2 on days 139 to 142 of gestation resulted in an average delivery day of 143 ± 1 compared with an average delivery day of 147 ± 1 in control ewes. The average gestation period in Rambouillet ewes typically falls around 147 days. Furthermore, lamb weight tended to be lower in the E group compared to the C group (4.12 ± 0.23 vs 4.77 ± 0.21 kg, P = 0.07). This observation can be rationalized as the induction of earlier delivery may result in slightly lower birth weights for the lambs. Also, the presence of both single and twin pregnancies included in the experiment can impact the birth weight of the lambs, as single pregnancies typically result in higher birth weights compared to twin pregnancies. No difference in lamb vigor was found between the E and C groups (2 ± 0.39 vs 1 ± 0.48, P = 0.13). Several factors may contribute to the lack of differences in lamb vigor. 1) The timing of lamb vigor assessment conducted a few hours after birth may have not captured initial differences that could have emerged in the following days. In fact, a couple of days later, two lambs in the E group experienced sudden death, while no death was reported in the lambs of the C group. 2) Despite the E group experiencing parturition around 10 days earlier than the C group, both groups were likely at a similar stage of lung maturity. This suggests that the timing of parturition alone may not determine lamb vigor.
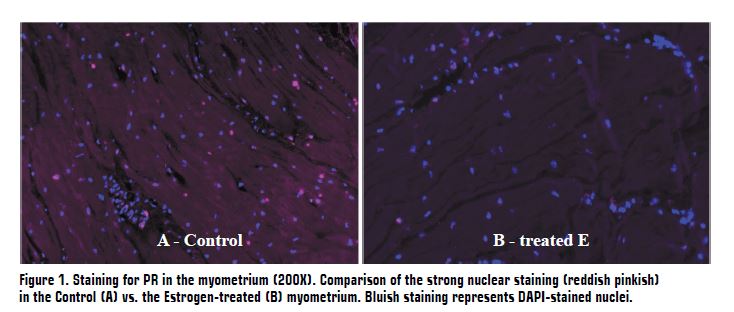
In experiment 2, a significant increase in E2 systemic levels was found in the E group compared with C group (149.21 ± 55.93 pg/mL vs 30.61 ± 11.73, P = 0.01), which suggests that E treatment effectively increased E2 levels. Estradiol has been related with the promotion of myometrial contractility and initiation of parturition. Together with the results in experiment 1, this finding suggests that a higher concentration of E2 could drive earlier delivery. A downregulation of myometrial P4 receptor protein expression was observed in the E group compared to C group (27.1 ± 36.7 vs 42.0 ± 3.68 intensity units, P = 0.01; Figure 1). This finding could have an important implication in the control of parturition, as P4 receptors play a crucial role in maintaining the inhibitory effects of P4 on uterine contractility promoting uterine quiescence during pregnancy. Therefore, the fact that the number of myometrial P4 receptors decreases with higher estrogen levels making the cells less responsive could potentially contribute to the initiation of parturition. No differences were found in P4 systemic plasma levels in E vs C (6.50 ± 1.42 vs 8.99 ± 1.42 ng/mL, P = 0.24), which together with the results in experiment 1, suggest that local changes in myometrial P4 receptors, rather than systemic P4 levels, might be more critical in regulating myometrial activation.
The findings of both experiments highlight the complex interplay between myometrial P4 receptor expression, systemic P4 and E2 concentrations, and the timing of parturition. Overall, these findings indicate that E2 concentrations have a notable effect on the timing of parturition and may contribute to slight differences in birth weight, but its impact on fetal maturation, as assessed by lamb vigor after birth, appears to be limited. Therefore, it can be concluded that E2 concentrations primarily affect the timing of parturition rather than directly influencing fetal maturation and that the downregulation of myometrial P4 receptor protein expression with the increase of E2 levels supports the important role of E2 in initiating labor. However, future research is needed to clarify the precise mechanism of parturition and fetal maturation to enhance reproductive management strategies in livestock.